在体育竞技的浩瀚星空中,人类对速度的追逐如同永不停歇的恒星,而男子100米短跑9.58秒的世界纪录,恰似其中最耀眼的超新星。这一由牙买加“闪电”尤塞恩·博尔特于2009年柏林世锦赛创造的数字,不仅是一串冰冷的计时,更是人类突破生理与心理边界的象征。它既是运动科学的里程碑,也是竞技精神的永恒丰碑。
一、从“不可能”到“新纪元”:百米纪录的百年突围

人类对百米速度的探索始于19世纪末的粗粝实验。1896年雅典奥运会上,美国选手托马斯·平克以11.8秒的“蹲踞式起跑”首开纪录时,或许难以想象百年后的赛道会进化到何种境地。这个在当时足以称雄的成绩,如今仅相当于中国国家二级运动员水平,折射出人类运动能力的几何级跃迁。
20世纪见证了速度探索的两次质变。1912年斯德哥尔摩奥运会上,唐纳德·利平科特以10.6秒首破11秒大关,这一被国际田联认证的首个正式世界纪录,标志着现代短跑技术的体系化开端。而1968年墨西哥奥运会的9.95秒,则粉碎了“人类无法突破10秒”的生物学诅咒——吉姆·海因斯冲线时的喃喃自语“上帝!这道门原来是虚掩的”,成为竞技史上最富哲学意味的注脚。
二、解剖“闪电”:9.58秒的生物学密码
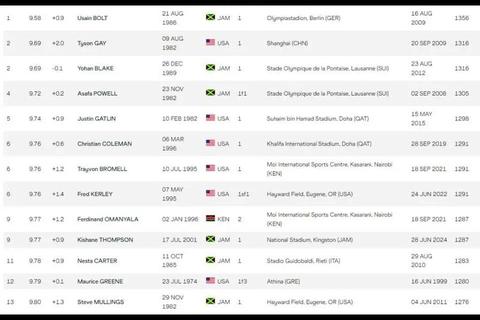
博尔特的身体构造堪称进化馈赠的杰作。1.96米身高带来的1.16米腿长,配合2.44米的惊人步幅,使他在41步内完成百米征途,这种“长程加速引擎”彻底颠覆了传统短跑选材理论。运动生物力学显示,其60-80米区间最高瞬时速度达44.64公里/小时,相当于城市道路限速标准,而支撑这种爆发力的腿部肌肉纤维强度达到常人两倍以上。
这种天赋并非孤立存在。神经肌肉协调性让他在起跑反应时达到0.146秒(柏林世锦赛数据),接近人体神经传导极限;乳酸代谢能力使肌肉在高强度收缩后仍保持弹性。更关键的是,博尔特将“后程加速”战术演绎到极致——在北京与柏林两届大赛中,后半程分别仅用4.65秒和4.71秒,这种“越跑越快”的反物理现象,重新定义了短跑的能量分配模型。
三、科技赋能:纪录背后的隐形推手
现代训练体系的多维革新,为突破速度壁垒提供了底层支撑。柏林之战中,博尔特的黄金钉鞋仅重96克,碳纤维底板与微型钉刺设计使触地时间缩短至0.08秒,能量损耗降低15%。高速摄像系统的帧级动作解析、风洞实验室的0.2米/秒风速控制、基于肌电信号的个性化力量训练,构成系统化的科技矩阵。
营养与康复医学的进步同样功不可没。实时血乳酸监测指导的能量补给策略、冷冻疗法维持的肌肉超量恢复、高压氧舱催生的细胞再生效率,这些原本属于航天医学的尖端技术,如今已成为顶级运动员的日常标配。数据显示,当代短跑选手的年训练损伤发生率较20年前下降37%,持续高强度输出的可能性提升2.3倍。
四、极限之壁:9.58秒的永恒挑战
生理学家通过肌肉力学模型推算,人类百米的理论极限约为9.45秒,但这仅是理想状态下的数学推演。现实中的多重屏障更为严峻:跟腱承受的峰值负荷达780公斤,相当于自身体重8倍;触地瞬间膝关节剪切力突破3000牛顿,接近骨骼强度阈值;心血管系统需在9秒内输送25升富氧血液,这对心室壁厚度与心率调节提出严苛要求。
心理层面的挑战同样不容小觑。诺亚·莱尔斯2024年巴黎奥运会的9.79秒虽未破纪录,但其赛后的技术分析显示,起跑阶段的0.4秒迟疑直接损失0.15秒优势。这种微观时间尺度的竞争,要求运动员在承受每秒11米风速、140分贝噪音和亿万目光注视下,仍保持神经系统的绝对冷静。
五、未来赛道:突破路径的多元可能
基因编辑技术或许将打开新的可能性。ACTN3基因(快肌纤维表达基因)的定向优化、ANGPTL3蛋白(能量代谢调节因子)的靶向增强,已在实验室环境中使小鼠的运动能力提升18%。尽管争议犹存,但科学界预测,基因改良运动员的出现可能将极限推至9.3秒区间。
训练理念的颠覆性创新正在酝酿。东京大学开发的“神经-肌肉耦合训练系统”,通过脑机接口直接优化运动指令传递效率;洛桑联邦理工学院的“仿生外骨骼”虽暂被国际田联禁用,但其辅助模式下已实现9.4秒模拟成绩。这些“科技兴奋剂”的边界,将成为体育界未来的核心争议。
永恒的竞速,不灭的火种
当博尔特在柏林夜空划出那道金色轨迹时,他不仅改写了计时器上的数字,更点燃了人类对自身潜能的永恒追问。从平克笨拙的皮鞋起跑,到碳纤维钉鞋的毫米级优化;从海因斯推开“虚掩之门”,到基因图谱揭示的速度密码,这场跨越三个世纪的竞速史诗,始终在证明:所谓极限,不过是等待突破的起点。在科技与天赋的交响中,下一个9.58秒的挑战者,或许正在某个训练场的起跑器上,蓄势待发。